




В лаборатории Б. Сигурдсона [7] были получены доказательства, что типичную медленную инфекцию овец – висну – вызывает вирус, оказавшийся по своим свойствам очень схожим с давно и хорошо известными онкорнавирусами. Понятно, что это открытие способствовало представлению о том, что все медленные инфекции вызываются вирусами. Укреплению этого мнения в большой мере способствовало и последующее установление вирусной этиологии известной еще с 1933 г. медленной инфекции детей и подростков – подострого склерозирующего панэнцефалита, – причиной которого, как оказалось, является вирус кори, возбудитель давно и хорошо известной детской заразной болезни [8].
Более того, в последующие годы был накоплен богатый фактический материал, прямо свидетельствующий о способности многих вирусов–возбудителей острых заразных заболеваний вызывать в организме человека или животных развитие медленной формы инфекционного процесса, полностью отвечавшего всем четырем признакам медленных инфекций. В числе таких возбудителей оказались вирусы кори, краснухи, герпеса, клещевого энцефалита, инфекционной анемии лошадей, гриппа, лимфоцитарного хориоменингита, бешенства, вирусы семейства папова, африканской лихорадки свиней, иммунодефицита человека и др. [9].
А между тем уже начиная с первых сообщений Б. Сигурдсона, подробно описавшего хорошо известную до того и распространенную болезнь овец – скрепи, – в литературе начали появляться сообщения, описывающие особую группу медленных инфекций человека и животных, патоморфологические изменения при которых в организме, как и при скрепи, отличались весьма существенным своеобразием: отсутствовали признаки воспаления и наряду с этим в центральной нервной системе развивалась картина выраженного первично–дегенеративного процесса в головном, а иногда и в спинном мозге. Изменения выражались в картине гибели нейронов, накоплении амилоидных бляшек и выраженном глиозе. В итоге все эти изменения приводили к формированию так называемого губкообразного состояния (status spongiosus) мозговой ткани (рис. 1), что и послужило основанием для обозначения этой группы заболеваний как «трансмиссивные губкообразные энцефалопатии» (ТГЭ) [10]. Именно трансмиссивность губкообразного состояния только мозговой ткани и служит патогномоничным признаком этих заболеваний.
Несмотря на точные доказательства инфекционной природы ТГЭ, на протяжении нескольких десятилетий не удавалось обнаружить возбудителей этих заболеваний. Вместе с тем накапливались данные, которые не прямо, но косвенно позволяли судить о некоторых свойствах предполагаемых возбудителей. Исследователи накопили большой фактический материал, разносторонне изучая инфицированную мозговую ткань. При этом оказалось, что предполагаемый инфекционный агент: проходит через бактериальные фильтры с диаметром пор от 25 до 50 нм; не размножается на искусственных питательных средах; воспроизводит феномен титрования; накапливается до концентрации 105–1011 ИД50 в 1 г мозговой ткани; способен к адаптации к новому хозяину, что нередко сопровождается укорочением инкубационного периода; способен первоначально репродуцироваться в селезенке и других органах ретикулоэндотелиальной системы, а затем и в мозговой ткани; обладает генетическим контролем чувствительности некоторых хозяев; имеет специфический для конкретного штамма круг хозяев; способен изменять патогенность и вирулентность для разного круга хозяев; селекционируется из штаммов дикого типа; воспроизводит феномен интерференции медленно накапливающегося в организме штамма с быстро накапливающимся; обладает способностью к персистенции в культуре клеток, полученных из органов и тканей зараженного животного [11,12].
Перечисленные признаки свидетельствовали о весьма большом сходстве их с признаками широко известных вирусов. Вместе с тем у предполагаемых возбудителей был обнаружен и ряд необычных характеристик. Возбудители ТГЭ оказались устойчивы к действию ультрафиолета, проникающей радиации, ДНКазы и РНКазы, ультразвука, глютаральдегида, b–пропиолактона, формальдегида, псораленов, толуола, ксилола, этанола, нагревания до 80 °С и даже неполностью инактивировались после кипячения [12].
Казалось совершенно естественным обозначить предполагаемых возбудителей ТГЭ как «необычные вирусы» или даже как «медленные вирусы» [10–12]. Однако вскоре эта неопределенность в обозначениях, а главное, в понимании природы возбудителей ТГЭ была устранена благодаря работам американского биохимика С. Прузинера. Им были использованы зараженные хомяки, в мозговой ткани которых накапливалось возбудителя в 100 раз больше, чем в мозговой ткани мышей. Получив мозговую ткань с высокой концентрацией возбудителя скрепи, С. Прузинер приступил к постепенной ее очистке, одновременно строго следя за сохранением инфекционных свойств. В результате такого подхода удалось установить безнуклеиновую, чисто белковую природу возбудителя: полученный инфекционный белок был представлен молекулами одного вида с молекулярной массой 27–30 кДа [13]. С. Прузинер предложил обозначить обнаруженный им инфекционный белок как «инфекционный прионный белок», а в качестве инфекционной единицы использовать термин «прион», т.е прион как инфекционная единица состоит из молекул инфекционного прионного белка [14].
Оказалось, что прионный белок может существовать в двух формах, т. е. белок того же аминокислотного состава и той же самой молекулярной массы обнаруживается в организме всех млекопитающих, включая человека, а его наивысшая концентрация определяется в нейронах. Учитывая его клеточное происхождение, этот прионный белок был назван «нормальным» или «клеточным прионным белком», обозначаемым символом PrPС (аббревиатура от английского – Prion Protein Cell) [15,16].
Синтез PrPC кодируется геном PRNP, расположенном в коротком плече хромосомы 20 у человека и хромосомы 2 у мыши. Ген является высококонсервативным, и наивысшие уровни его экпрессии зафиксированы в нейронах, где концентрация иРНК для РrPC в 50 раз выше, чем в клетках глии [16,17].
Оказалось, что клеточный прионный белок РrPC играет важную роль в жизнедеятельности организма млекопитающих: он участвует в передаче нервных импульсов между окончаниями нервных волокон, способствует сохранению устойчивости нейронов и клеток глии к окислительному стрессу, вовлекается в процессы регуляции содержания внутриклеточного кальция (Са2+) в нейронах, но самое главное – поддерживает циркадные (от лат. circa – около и dies – день), т.е. околосуточные, ритмы активности и покоя в клетках, тканях, органах и в организме в целом [18,19].
Дополнительным доказательством подобной роли клеточных прионов послужило открытие в 1986 г. Логарези и соавт. новой медленной инфекции, связанной со снижением в организме синтеза клеточного прионного белка. Такие пациенты начинали страдать резким сокращением продолжительности сна, галлюцинациями, утратой циркадных ритмов и деменцией, а затем и вовсе погибали от бессоницы. Именно поэтому болезнь получила название «смертельная семейная бессоница» [10].
В организме людей и животных, страдающих ТГЭ, прионный белок обнаруживается в другой форме, обозначаемой как PrPSc. Предложенная аббревиатура основана на том, что природным резервуаром инфекционного прионного белка является организм овец и коз, у которых спонтанно может развиваться уже упомянутое выше заболевание скрепи (от англ. Scrapie) [20].
Сегодня известно, что процесс накопления инфекционных прионных молекул, т.е. воспроизведение себе подобных, осуществляется благодаря изменениям третичной стуктуры в белковой молекуле клеточного прионного белка PrPC, суть которых выражается в превращении части a–спиральных доменов в b–вытянутые тяжи [21]. Этот процесс превращения нормального клеточного белка в инфекционный носит название конформационного, т.е. связанного только с изменением пространственной структуры молекулы белка, но не его аминокислотного состава (рис. 2).
Схема процесса накопления молекул
инфекционного прионного
белка PrPSc
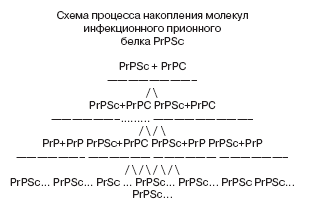
Таким образом, процесс накопления инфекционного прионного белка PrPSc в зараженном организме происходит не за счет синтеза таких молекул de novo, а за счет конформационных изменений уже существущих и предварительно синтезирующихся вновь молекул клеточного белка PrPC. Этот процесс носит лавинообразный характер (рис. 2) [22,23].
Медленные инфекции, вызываемые прионами, получили наименование «прионные болезни», что с тех пор широко используется в научной литературе наряду с существовавшим ранее и использующимся до сих пор определением – «трансмиссивные губкообразные энцефалопатии» [24].
Открытие прионов послужило мощным стимулом для успешной расшифровки этиологии ряда нейродегенеративных заболеваний, в патогенезе которых уже давно было описано формирование губкообразной энцефалопатии. Среди таких заболеваний следует прежде всего выделить болезнь Крейтцфельда–Якоба (БКЯ), хорошо известную еще с 1920–х гг. Болезнь встречается с частотой 1 случай на 1 млн населения в год. Однако существуют кластеры (в Чили, Израиле, Словакии), где заболеваемость оказывается значительно выше [25].
Клиническая картина БКЯ включает в себя быстро прогрессирующее слабоумие, миоклонус и заметно прогрессирующие двигательные нарушения, что, как правило, в течение нескольких месяцев приводит к смертельному исходу [10]. В отличие от всех известных инфекционных болезней, для БКЯ характерно, что из 100% заболевающих в 85% случаев БКЯ возникает как спорадическое заболевание, как правило, у людей в возрасте 40–69 лет, в 10–15% случаев – как наследственное заболевание, и менее чем в 1–5% случаев эта болезнь развивается как инфекционная, т.е. в результате типичного внешнего заражения. Последний вариант подразумевает так называемые ятрогенные случаи, связанные с медицинскими манипуляциями или пересадками тканевых материалов от человека человеку (табл. 1) [10,25].
Из данных таблицы 1 следует, что все случаи ятрогенной передачи БКЯ связаны либо с пересадками и использованием биологических материалов, полученных от находящихся в инкубационном периоде скрыто инфицированных людей, либо в результате недостаточной обработки инструментария, учитывая весьма высокую степень термостабильности инфекционного прионного белка PrPSc.
Указанные случаи вызвали необходимость неоднократных выступлений ведущих специалистов мира с обращениями к медицинской общественности, в которых рекомендовалось: инструменты для хирургических процедур в неврологических клиниках или отделениях больниц стерилизовать автоклавированием в течение 1 ч при температуре 135 °С; пластиковые, металлические и прочие предметы (столы, тазы, полы, стены и др.) обрабатывать в течение 1 ч 0,5%–ным раствором хлорноватистокислого натрия или 1N раствором гидрата окиси натрия (NaOH); руки следует обрабатывать в течение 10 мин. 1N раствором гидрата окиси натрия с последующим тщательным мытьем с мылом [26].
Помимо уже упомянутой выше скрепи овец и коз, инфекционный прионный белок оказался причиной трасмиссивной энцефалопатии норок, хронической изнуряющей болезни некоторых видов оленей и лосей, однако в конце 1980–х гг. к ним прибавились еще три прионных болезни животных, что было связано с разразившейся в 1986 г. в Великобритании эпизоотией трансмиссивной губкообразной энцефалопатии крупного рогатого скота (ТГЭКРС). Это заболевание было обусловлено массовым заражением молодняка, при выкармливании которого, как оказалось, использовали зараженную инфекционным прионным белком мясокостную муку.
Напомним, что ранее мясокостная мука широко использовалась во многих странах при выкармливании телят, а также в бытовых условиях, например, при выкармливании щенков, что в последнем случае считалось более удобным вместо использования аптечного глюконата кальция.
Сырьем для мясокостной муки служат скелеты крупного и мелкого рогатого скота и другие, не идущие в пищу человека части туш коров и овец.
В технологию получения такой муки после процессов тщательного измельчения исходного сырья включена также обработка активными жирорастворителями и, наконец, термообработка при температуре 130 °С.
Однако в конце 70–х гг. ХХ в. в Великобритании предприниматели решили повысить питательную ценность мясокостной муки, снизив для этого режим термообработки до 110 °С, а также уменьшив количество веществ, экстрагирующих жир. Именно эти изменения технологии и сыграли роковую роль в развитии эпизоотии крупного рогатого скота [10].
Эпизоотия развивалась с нарастающей силой. Болезнь охватила практически все графства, число заболевших коров постоянно увеличивалось и в 1992 г. достигло пика, когда каждую неделю регистрировали до 1 тыс. случаев заболеваний.
Клинически болезнь проявлялась прежде всего, как выражаются ветеринары, в «потере животным кондиции» и снижении удойности. У коров и быков уменьшается время жевания пищи, они становятся особо чувствительными к прикосновению и звуку, отмечается ненормальная осанка – голова низко опущена, нарушаются движения, животное склонно к падениям. С самого начала были подмечены нарушения психики: у больных животных появляется страх, боязнь дверных проемов и наряду с этим появляются приступы бешенства («болезнь бешеной коровы») [27].
Так же как у овец и коз при скрепи, у погибших коров при вскрытии в головном и иногда в спинном мозге обнаруживается гибель нейронов, образование вакуолей, формирование на этой основе губкообразного состояния мозговой ткани и развитие глиоза. На гистологических срезах ткани головного мозга обнаруживается присутствие инфекционного прионного белка PrPSc в области вакуолизации и на поверхности еще сохранившихся нейронов.
За 10 лет в Великобритании погибло около 200 тыс. голов крупного рогатого скота, хотя правительством Великобритании начиная с 1988 г. были предприняты энергичные меры: с июля 1988 г. введен запрет на кормление жвачных животных белковосодержащими кормами, приготовленными из органов и тканей жвачных животных; с августа того же года все животные с подозрением на губкообразную энцефалопатию коров подлежали убою, а затем и уничтожению в электрических печах [10,27].
Начиная с 1993 г. эпизоотия пошла на убыль, однако в результате число известных ранее трансмиссивных губкообразных энцефалопатий удвоилось: к известным ранее скрепи, трансмиссивной энцефалопатии норок и изнуряющей хронической болезней оленей и лосей прибавились собственно ТГЭКРС, трансмиссивная губкообразная энцефалопатия кошек и трансмиссивная губкообразная энцефалопатия экзотических копытных.
Заболевание сначала бродячих, а затем и домашних кошек было обусловлено употреблением ими в пищу инфицированных мяса и внутренностей больных и погибших коров и быков (табл. 2).
На этом основании Европейский союз запретил около 400 лекарственных препаратов, для приготовления которых использовались органы и ткани жвачных.
Случаи губкообразной энцефалопатии возникли и у находящихся в Лондонском зоопарке пумы, двух гепардов, оцелота и тигра. Причина проста – всех их в течение длительного времени кормили расколотыми головами и мясом крупного рогатого скота [27].
Губкообразная энцефалопатия экзотических копытных описана у 5 видов диких животных: сернобыка, аравийского орикса, канны, сахарского орикса и большого куду. Семнадцать случаев заболеваний среди этих животных в зоопарках также обусловлены использованием при их выкармливании мясокостной муки.
Прионные болезни крупного рогатого скота, помимо Великобритании, вскоре были зарегистрированы вo Франции, Германии, Швейцарии, Италии, Португалии, Ирландии, Нидерландах, Дании, США, Омане, Китае и на Фолклендских островах. К сожалению, с годами список стран, зарегистрировавших у себя случаи ТГЭКРС, постепенно увеличивался и к 2006 г. достиг уже 40, что было связано либо с импортом уже зараженных животных либо с использованием в этих странах инфицированной мясокостной муки. Столь широкое распространение этой прионной болезни крупного рогатого скота естественно обосновывало беспокойство по поводу возможности передачи заболевания от животных человеку, тем более, что был уже твердо установлен путь передачи инфекционного агента с зараженой пищей.
В марте 1996 г. это беспокойство было значительно подкреплено сообщением, переданным Великобританией в ВОЗ, о 10 случаях смерти молодых людей от так называемого нового варианта БКЯ (нвБКЯ). Через месяц о таком же случае сообщила Франция, а через год на территории Великобритании были выявлены еще 5 случаев этой болезни. К 1998 г. в мире стало известно о 24 случаях, а к настоящему времени их число в Европе уже достигло 200 [10,27].
В самом начале было подмечено, что нвБКЯ отличается от классической БКЯ рядом характерных симптомов: болезнь поражает молодых людей в возрасте в среднем до 30 лет, среди симптомов отмечают развитие тревожного состояния, бессоницу, хорею, миоклонус и прогрессирующую атаксию. Патоморфологическая картина, хотя и была характерна для ТГЭ, однако отличалась обязательным присутствием больших амилоидных бляшек в мозжечке и в коре головного мозга, окруженных многочисленными вакуолями [10,27].
Учитывая доказанность алиментарного пути заражения крупного рогатого скота с одной стороны, и возможность для инфекционного прионного белка преодолевать межвидовые барьеры с другой, подозрение, естественно, падало на вероятность развития нвБКЯ в связи с употреблением в пищу зараженных продуктов питания.
Для выяснения причин на мышах было проведено сравнительное исследование трех штаммов инфекционного прионного белка PrPSc, выделенного: 1) из мозговой ткани коровы, погибшей от ТГЭ, 2) из мозговой ткани овцы, погибшей от скрепи, и 3) из мозговой ткани молодого человека, погибшего от нвБКЯ. Результаты исследования по трем генетическим маркерам – продолжительность инкубационного периода, скорость гибели зараженных мышей и профиль поражения ЦНС – подтвердили сходство прионов, выделенных от нвБКЯ, с прионами, выделенными именно только от коровы, погибшей от ТГЭ [28,29].
Исследования, проведенные на трансгенных мышах, показали, что степень чувствительности организмов к инфекционному прионному белку (PrPSc) доноров определяется структурной близостью к нему клеточного прионного белка (PrPC) реципиента [23]. Однако такая закономерность оказывается не абсолютной, а является лишь тенденцией, т.к. все приведенные выше примеры, и главный из них – эпизоотия ТГЭКРС с ее последствиями для других видов животных, – прямо свидетельствуют о том, что увеличение дозы инфекционного материала и частоты его введения в организм способствуют успешному преодолению барьеров нечувствительности организма к инфекционному прионному белку.
Кроме этого, следует особо подчеркнуть, что в изучении природы чувствительности и нечувствительности к прионным болезням большую роль играет генотип конкретного животного или конкретного человека. И этот генетический контроль обусловлен видом мутаций, происходящих в гене PRNP, кодирующем синтез клеточного прионного белка PrPC.
Сегодня картированы более 15 хорошо изученных мутаций в гене PRNP. Так, при случаях спорадической БКЯ у пациентов обнаружена мутация в кодоне 178, при которой происходит замена аспарагиновой кислоты на аспарагин. Но если при этом 129–й кодон кодирует валин, то действительно развивается БКЯ, а если же в положении 129 находится метионин, то развивается семейная смертельная бессоница. При мутации Про102Лей возникает синдром Герстманна – Штреусслера – Шейнкера [27].
Первые же попытки иммунодиагностики, иммунотерапии или иммунопрофилактики прионных болезней столкнулись с фактом полного отсутствия специфических антител в зараженном организме [10,24]. Подобный результат становится понятным, учитывая структурную близость инфекционного прионного белка PrPSc и его клеточной изофориммы PrPС, в связи с чем организм «рассматривает» белок PrPSc как «свой». Поэтому неудивительно, что испытания большого числа иммуномодуляторов с целью повлиять на изменение чувствительности мышей окончились неудачей, и только преднизолон, арахисовое масло и полианионы заметно снижали чувствительность мышей к возбудителю скрепи.
Клиническая диагностика прионных болезней основывается на уже описанной выше симптоматике. Лабораторная диагностика этих заболеваний включает в себя прямые и непрямые методы [30]. К первым из них относятся: электронно–микроскопическое определение скрепиассоциированных фибрилл, определяемых в инфицированных материалах или отпечатках; иммуноблоттинг с использованием моноклональных антител; метод пептидных зондов, основанный на использовании меченых синтетических пептидов, аминокислотная последовательность которых позволяет им связываться со структурой прионного белка.
Непрямые методы включают обычную гистологическую технику, благодаря которой в образцах биопсийного или аутопсийного материала определяют глиоз, формирование губкообразного состояния мозговой ткани и накопление в ней амилоидных бляшек; гистохимический метод основывается на выявлении с помощью красителей (конго красный, теофлавин Т) скоплений амилоида; биологический метод включает заражение испытуемым материалом лабораторных животных (биопроба), что, однако, связано с длительностью наблюдения, или заражением клеточных культур. Недавно для последнего варианта была предложена культура клеток N2a, заражение которой испытуемым материалом позволяет в 10 раз быстрее, чем с помощью биопробы, получить результат при сохранении того же уровня чувствительности [30].
Профилактика прионных болезней, помимо перечисленных выше мероприятий, основывается также на недопущении в пищу инфицированных мясных продуктов или других продуктов убоя. Именно в этой связи еще в 2000 г. главным государственным санитарным врачом РФ было выпущено постановление № 15 «О мерах по предупреждению распространения болезни Крейтцфельда–Якоба на территории Российской Федерации», предписывающее, в частности, «не допускать закупок мяса и мясных и других продуктов убоя крупного рогатого скота, а также получения их по гуманитарной помощи без предоставления документов, подтверждающих отсутствие в стране–экспортере заболеваний губкообразной энцефалопатии коров». Предписывалось также усилить контроль на местах за продажей мяса и мясных продуктов [27].
Сегодня надо признать, что проблема лечения прионных болезней остается нерешенной и все многочисленные попытки применения в клинической практике антибиотиков, ацикловира, стероидов или тиамина при БКЯ неизменно заканчивались неудачей [26,27]. Поэтому, мы вправе пока говорить лишь о перспективных препаратах, дающих основание для дальнейших исследований. К таким соединениям уже сегодня относятся: группа сульфатированных глюкозоаминогликанов, включающая пентозан полисульфат, каррагенан, декстрансульфат 500; полиеновый антибиотик амфотерицин В и его производные, антибиотик антрациклин, порфирины и фталоцианины, полиамины и некоторые другие.
Все возрастающий мировой интерес к проблеме прионов и прионных болезней связан не только с продолжающимся увеличением числа случаев заболеваний нвБКЯ людей и случаев заболевания ТГЭКРС, но и с абсолютно новыми качествами самого возбудителя этих заболеваний.
Сегодня уже накапливаются данные о способности некоторых белков к изменению своей третичной и вторичной структуры даже без мутационных изменений, что свидетельствует об эссенциальной способности таких белков к патологической конформации [27]. Добавим, что появление других заболеваний, основанных на нарушениях третичной или вторичной структуры белков, позволило подразделить их на инфекционные и неинфекционные амилоидозы и ввести новое определение – «конформационные болезни» [31]. Есть основания полагать, что такие конформационые белки могут выполнять роль главных регуляторов в организме, в том числе и такого важнейшего процесса, как ограничение ими самой продолжительности жизни.



![Таблица 2. Инфицированность различных органов и тканей жвачных на примере заболевших овец и коз [27]](/data/articles/Image/t18/n6/381-4.gif)
Литература
1. Sigurdsson B. Brit. Vet. J. 1954; 110: 255–270.
2. Idem Ibid: 307–322/
3. Ibem Ibid: 341–354/
4. Gajdusek D.C., Zigas V. New Engl. J. Med. 1957; 257: 974–978.
5. Gajdusek D.C., Gibbs C.J. Nature 1966; 209: 794–796.
6. Hadlow W.J. Lancet 1959; 2: 289–290.
7. Sigurdsson B., Thormar H., Palsson P.A. Arch. Ges. Virusforsch. 1960; 10: 368–381.
8. Horta–Barbosa L., Hamilton R., Writting B. et al. Nature 1969; 221: 974.
9. Зуев В.А. Медленные вирусные инфекции человеки животных.М., 1988.
10. Зуев В.А., Завалишин И. А., Ройхель В.М. Прионные болезни человека и животных (руководство для врачей). М., 1999.
11. Gajdusek D.C. Virology (Ed. B.N. Fields) New York, 1985; 1519–1557.
12. Gajdusek D.C. Subviral Pathogenesis of Plants and Animals:Viroids and Prions. New York 1985; 483–544.
13. Prusiner S.B., McKinley M.P., Groth D.F. et al. Proc. Natl. Acad. Sci. USA 1981; 78: 6675–6679.
14. Prusiner S.B. Science 1991; 252: 1515–1522.
15. Weissmann C. Nature 1991; 352: 679–683.
16. Bolton D.C., McKinley M.P., Prusiner S.B. Science 1982; 218: 1309–1311.
17. McKinley M.P., Bolton D.C., Prusiner S.B. Cell 1983; 35: 57–62.
18. Tobler J., Gaus T., Deboer P. et al. Nature 1996; 380: 639–642.
19. Harris D.A., Falls D.L., Johnson F.A. et al. Proc. Natl. Acad. Sci. USA 1991; 88: 7664–7668.
20. Oesch B., Westaway D., Walchli M., et al. Cell 1985; 40: 735–746.
21. Prusiner S.B. Neurodegenerative Diseases (eds. G. Golls & J.M. Stutzmann) Acad. Press 1996; 23–80.
22. Prusiner S.B. Prions Prions Prions (ed. S.B. Prusiner) Berlin 1998; 1–18.
23. Huang Z., Prusiner S.B., Cohen F.E. Prions Prions Prions (ed. S.B. Prusiner) Berlin 1998; 49–63.
24. Palmer M., Collinge J., Prion Diseases (eds. J. Collinge& M. Palmer) Oxford 1997; 1–17.
25. Collinge J., Palmer M. Prion Diseases (eds. J. Collinge & M.S. Palmer) Oxford 1997; 18–55.
26. Brown H. Antiviral Chem. Chemother. 1990; 1: 75–83.
27. Покровский В.И., Киселев О.И., Черкасский Б.Л. Прионы и прионные болезни. М., 2004.
28. Bruce M.E., Will R.G., Ironside J.M. et al. Nature 1997; 389: 498–501.
29. Hill A.F., Desbruslais M., Joiner S. et al. Ibid.: 448–450.
30. Григорьев В.Б., Покидышев А.Н., Кальнов С.Л., Клименко С.М. Вопр. вирусол. 2009; 5: 4–9.
31. Carrell R.W., Lomas D.A. Lancet 1997; 350: 134–138



