




FGU “MNII GB named after Gelmgoltsa Rosmedbiotechnology”
Moscow
Review is devoted to sources of origin of oscillatory potentials, their role in diagnostics of various eye diseases and according to particular retinal structures.
Впервые осцилляторные потенциалы (ОП) были описаны как группа осцилляции на восходящей части b–волны электроретинограммы (ЭРГ) лягушки исследователями Granit и Munsterhjelm в 1937 году. В 1953 году Cobb и Morton зарегистрировали около 4–6 осцилляций в ЭРГ у человека.
В 1962 году Yonemura с соавторами предложили термин «осцилляторные потенциалы». В 1972 году Wachtmeister показал, что для получения наилучших результатов регистрации ОП необходимы особые условия адаптации – мезопические, при которых функционируют обе системы: палочковая и колбочковая. В то же время авторами Wachtmeister и Algvere в 1972 году была отмечена необходимость выделения ОП путем фильтрации ЭРГ на частотах около 80 и 160 Гц [1].
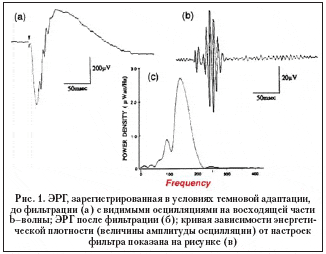
ОП – это волны (осцилляции) на восходящей части b–волны, регистрируемые при записи ЭРГ. Согласно общепринятым стандартам (ISCEV, 2004 год) их выделяют при темновой или световой адаптации при расширенном зрачке, используя стандартный белый стимул (1,5–3,0 фотопические кд•с/м2), предъявляемый с интервалом через каждые 15 с при темновой и 1,5 с при световой адаптации с настроенным фильтром пропускания частот на диапазон от 75–100 Гц до 300 Гц и выше. При этом в анализе используются только вторая и последующие волновые последовательности в связи с возможным влиянием артефактов и частым искажением первого сигнала [2,3,10].
На рисунке 1 [2] представлена записываемая кривая ЭРГ при указанных выше условиях (при темновой адаптации).
Исследование происхождения ОП у теплокровных животных показало, что наибольшая амплитуда ОП регистрируется во внутренних слоях сетчатки [8,9]. Клеточные структуры в более глубоких слоях сетчатки – фоторецепторы, горизонтальные клетки – не принимают участие в формировании ОП, одной из возможных причин этого являются, например, частотные характеристики горизонтальных клеток, которые слишком медленны для генерации ОП. Пигментный эпителий и клетки Мюллера не принимают участия в генерации ОП, доказательством чего является регистрация ОП в сетчатке, изолированной от пигментного эпителия [11]. Исследования указывают на то, что источником ОП являются внутренние слои сетчатки, а точнее, в большей степени электрохимические процессы в аксонных терминалях биполярных клеток, в отростках амакриновых клеток, интерплексиформных клетках и дендритах ганглиозных клеток [1].
Полностью исчезают ОП при сохранности рецепторного потенциала после использования аспартата или глютамата, которые блокируют синаптическую фоторецепторную передачу. Наибольшую роль в генерации ОП играют нейрональные взаимодействия, при этом не исключается, что мембранные процессы, происходящие в клетках, играют одну из ведущих ролей в происхождении ОП [6,7]. Имеются данные о регистрации ОП от деполяризующихся амакриновых клеток. Предполагается, что гиперполяризующиеся амакриновые клетки не продуцируют ОП на включение стимула, но ОП могут возникать при его выключении. При регистрации ЭРГ после введения глицина в стекловидное тело глаза кролика ОП исчезают. Так как глицин является веществом, которое вызывает морфологические изменения большинства амакриновых клеток, то предполагается, что одним из источников ОП являются амакриновые клетки [12]. Интерплексиформные клетки также не могут быть исключены, как источник ОП. Эти нейроны обеспечивают центробежный путь в сетчатке путем создания пре– и постсинаптических контактов с амакриновыми клетками во внутреннем плексиформном слое, что показано в эксперименте у макаки. Они имеют пресинаптические контакты с биполярными и горизонтальными клетками в наружном плексиформном слое [17,18]. В единичных работах представлены результаты внутриклеточного отведения ОП от интерплексиформных клеток, в том числе деполяризующихся горизонтальных клеток. В этих же работах указывается на то, что ОП могут быть использованы как селективный зонд определения сохранности нейрональных связей в проксимальной сетчатке [12,17].
Было выявлено уменьшение осцилляторного ответа на введение каиновой кислоты, вызывающей набухание дендритов крупных ганглиозных клеток у кошки, что определяет ганглиозные клетки как источник ОП [1]. Однако при атрофии зрительного нерва амплитуда ОП практически не изменяется [13,14]. Объяснение этого, возможно, связано с местом генерации ОП в тормозном реципрокном соединении аксона амакриновой клетки с соседней ганглиозной клеткой. Поэтому вопрос о вкладе ганглиозных клеток в генерацию ОП остается открытым [1].
Изучение источников ОП по амплитуде осцилляций и количеству одновременно возникающих осцилляций в данной конкретной области (по «плотности источников тока») в сетчатке приматов in vivo показало, что наиболее вероятным источником ОП являются аксонные терминали биполярных клеток во внутреннем слое сетчатки. Также было показано, что источники a– и b–волн сетчатки находятся более дистально от ее внутреннего слоя [15,16].
ОП отображают активность нейронов, к которым стекается информация от всей сетчатки (пространственная интеграция). В светоадаптированных условиях пространственная интеграция уменьшается по сравнению с тем, что происходит в темноадаптированных условиях [19]. По мнению некоторых авторов, которые исследовали ОП в клинике, ОП являются индикатором ишемии и метаболических нарушений сетчатки центральных отделов глазного дна [Yonemura и др. 1962; Algvere, 1968; Algvere и Wachtmeister, 1972; Galloway и др., 1972; Miyake, 1990].
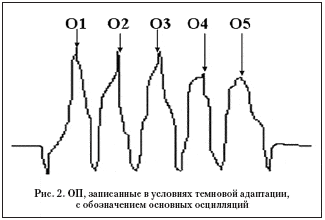
Среди регистрируемых основных пяти (O1–O5) волн осцилляторных потенциалов выделяют ранние осцилляции (волны O1–O3) и поздние осцилляции (O4–O5), рис. 2 [1].
Ранние ОП происходят около внутренних плексиформных слоев, в то время как поздние ОП возникают более дистально – во внутреннем ядерном слое, однако при этом не исключается их происхождение в проксимальных нейронах сетчатки. Различная амплитуда ОП является следствием сложных биоэлектрических процессов на клеточном уровне. Клиническим подтверждением этого положения является подавление или исчезновение ОП при окклюзии центральной артерии сетчатки, которая питает внутренние и средние слои сетчатки.
Ранние осцилляции преимущественно блокируются галоперидолом (блокатором допамина), ГАМК в насыщающих концентрациях или блокаторами ГАМК (пикротоксин или бикукулин). Амплитуда поздних осцилляций избирательно уменьшается под воздействием глицина [Korol и др., 1975, Wachtmeister, 1980] [1]. ГАМК и глицин – это аминокислоты, которые являются ингибиторными нейротрансмиттерами. Показано в эксперименте у собаки, что деполяризующиеся биполярные и амакриновые клетки – ГАМК–чувствительные, а глицин, выделяемый гиперполяризующимися биполярами, участвует в передаче информации по off–путям [Cunningham и Miller, 1980] [1]. Ингибиторные пути во внутреннем слое сетчатки, относящиеся к тормозным обратным связям в соединениях биполяров и амакриновых клеток у лягушки, и интерплексиформные клетки у кошки являются ГАМК–ергическими [Nakamura и др., 1979, Burchardt, 1972] [1]. С другой стороны, глицин избирательно уменьшает off–деполяризацию on–, off–клеток и определенных off–путей сетчатки карпа, не влияя на on–деполяризацию [Negishi и др., 1978] [1]. Таким образом, ранние ОП отражают состояние ингибиторных реципрокных взаимодействий, реализуемых в основном амакриновыми клетками on–путей сетчатки, а поздние ОП – off–путей внутреннего слоя сетчатки [1].
ОП не изменяются под воздействием таких возбуждающих нейротрансмиттеров, как ацетилхолин, карбахолин и мало меняются при действии других возбуждающих аминокислот, но заметно меняются под действием тормозных медиаторов, что указывает на то, что ОП отражают преимущественно нейрональную активность ингибиторных путей внутреннего слоя сетчатки [1].
Ранние ОП достоверно изменяются при предъявлении стимула с увеличивающейся частотой и значительно уменьшаются при частоте, превышающей порог восприятия палочек (20 Гц). Эти данные указывают на связь ранних осцилляций с палочковой системой сетчатки, так как порог колбочковой системы выше 20 Гц [20]. При ретинопатиях, первично поражающих палочковую систему сетчатки, ранние осцилляции уменьшались или исчезали, тогда как при поражении колбочковой системы изолированно уменьшались поздние ОП [1].
Было установлено при биохимических, гистологических и электрофизиологических исследованиях, что среди получаемых основных пяти (O1–O5) волн осцилляторных потенциалов ранние осцилляции (волны O1–O3) связаны с on–палочковыми ингибиторными путями передачи импульсов, в большей степени ГАМК–ергических (возвратные – feed–back), а поздние (O4–O5) – с off–колбочковыми ингибиторными возвратными глицинергическими путями передачи импульсов [1].
Учитывая сведения о происхождении ОП, была исследована их роль при различных заболеваниях глаз. Так, изменения ОП обнаружены при глаукоме, мягких экссудатах и неоваскуляризации при диабетической ретинопатии, что связано с уменьшением или прекращением кровотока по сосудам сетчатки и нарастанием ишемии ее внутренних слоев [1].
При окклюзии центральной артерии сетчатки или ее ветви прогноз исхода заболевания тем лучше, чем выше амплитуда ОП, что коррелирует с глубиной ишемии сетчатки; сходная зависимость наблюдается при нарушениях венозного оттока сетчатки [1]. При артериальной гипертензии и хронической почечной недостаточности амплитуда ОП также снижается, особенно поздних компонентов, что указывает на большую выраженность поражения колбочковых off–путей макулярной области при данной патологии [1].
Таким образом, источником ОП являются различные клеточные элементы сетчатки, исследование которых должно быть продолжено, так как многие вопросы их происхождения остаются невыясненными, несмотря на большое количество работ, посвященных ОП.
Очевидно, что при различных заболеваниях ОП изменяются в соответствии со степенью нарушения функций структур сетчатки, деятельность которых они отражают. Можно предположить, что дальнейшее изучение этой проблемы позволит представить новые патогенетические звенья многих заболеваний сетчатки и даст возможность разработать эффективные патогенетически обоснованные методы их диагностики и лечения.
Литература
1. Wachtmeister L. Oscillatory Potentials in the Retina: what do they Reveal.// Progress in retinal and Eye Research. –1998.– Vol. 17, No.4, pp.485 to 521.
2. Kolb H., Fernandez E., Nelson R. WEBVISION: The Organization of the Retina and Visual System – John Moran Eye Center, University of Utah, 2004.
3. Шамшинова А.М., Волков В.В. Функциональные методы исследования в офтальмологии. – М., Медицина, 2004.
4. Шамшинова А.М. Клиническая физиология зрения. – М., Научно–медицинская фирма МБН, 2006.
5. Kenyon GT, Moore B, Jeffs J, Denning KS, Stephens GJ, Travis BJ, George JS, Theiler J, Marshak DW.A model of high–frequency oscillatory potentials in retinal ganglion cells.Vis Neurosci. 2003 Sep–Oct;20(5):465–80.
6. Bui B.V., Armitage J.A. et al. Extraction and modeling of oscillatory potentials // Doc. Ophtalmol. – 2002. – Vol. 104, N1. – P. 17–36.
7. Janaky M., Goupland S.G., Benedek G. Human oscillatory potentials: components of rod origin // Ophthalmologica. – 1996. – Vol. 210, N6. – P. 315–318.
8. Ogden T.E. (1973) The oscillatory waves of the primate electroretinogram. Vision Res. 13, 1059–1074.
9. Ogden T.E. and Wylie R. (1971) Avian retina. I. microelectrode depth and marking studies of local ERG. J. Neurophysiol. 34, 357–361.
10. Michael F. Marmor, Graham E. Holder, Mathias W. Seeliger, Shuichi Y. Standard for clinical electroretinography.//Documenta Ophthalmologica. –2004.– 108: 107–114.
11. Foerster W.H., van de Grind W.A., and Gruesser O.J. (1977) Frequency transfer properties of three distinct types of cat horizontal cells. Exp. Brain Res. 29, 347–366.
12. Korol S., Leuenberger P.M., Englert U. and Babel J. (1975) In vivo effects of glycine on retinal ultrastructure and averaged electroretinogram. Brain Res. 97, 235–251.
13. Wanger P. and Persson H. (1983) Pattern–reversal electroretinograms in unilateral glaucoma. Invest. Ophtalmol. Vis. Sci. 24, 749–753.
14. Wachtmeister L. and el Azazi M. (1985) The oscillatory potentials of the electroretinogram in patients with unilateral optic atrophy. Ophthalmologica 191, 39–50.
15. Nelson R.A., Kolb H., Famiglietti E. and Gouras P. (1976) Neuronal responses in the rod and cone system of the cat retina: intracellular recordings and procain stains. Invest. Ophthalmol. Vis. Sci. 15, 946–953.
16. Smith E., Harwerth R., Crawford M.L.J. and Duncan G. (1989) Contribution on the retinal on–channels to scotopic and photopic spectral sensitivity. Visual Neuroscience 3, 225–239.
17. Boycott B.B., Dowling J.E., Fisher S.K., Kolb H.K. and Laties A.M. (1975) Interplexiform cells of the mammalian retina and their comparison with catecholamine containing retinal cells. Proc. R. Soc. Lond. 191, 353–368.
18. Dowling J.E. and Ehinger B. (1975) Synaptic organization of the dopamine containing interplexiform cells of the goldfish and Cebus monkey retina. Science. 188, 270–273.
19. Wachtmeister L. and Hahn I. (1987) Spatial properties of the oscillatory potentials of the electroretinogram in relation to state of adaptation. Acta Ophtalmol. (Copenh.) 65, 724–730.
20. Lachapelle P., Little J. and Polomeno R. (1983) The Photopic Electroretinogram in Congenital Stationary Night Blindness with Myopia. Invest. Ophthalmol. Vis. Sci. 24, 442–450.
21. Kazaryan A. A., Shamshinova A. M. Oscillatory potentials and ischemic processes in glaucoma//Annals of Ophthalmology. – 2006.– №4.
22. А.А. Казарян, О.Г. Пантелеева, А.М. Шамшинова, Ф.Г. Сеидова Нейрональные взаимоотношения в сетчатке и глаукома// ГЛАУКОМА: Всероссийская научно–практическая конференция, Сборник трудов конференции, МНИИ глазных болезней им. Гельмгольца. – 2004.



